
Vestiges of a Beginning: Clues to the Emergent Biosphere Recorded in the Oldest Known Sedimentary Rocks
Stephen J. Mojzsis, T. Mark Harrison, Department of Earth and Space Sciences and IGPP Center for Astrobiology, University of California, Los Angeles, CA 90095-1567, USA
published in GSA Today, April 2000
ABSTRACT
Recent explorations of the oldest known rocks of marine sedimentary origin from the southwestern coast of Greenland suggest that they preserve a biogeochemical record of early life. On the basis of the age of these rocks, the emergence of the biosphere appears to overlap with a period of intense global bombardment. This finding could also be consistent with evidence from molecular biology that places the ancestry of primitive bacteria living in extreme thermal environments near the last common ancestor of all known life. To make new advances in understanding the physical, chemical, and biological states of early environments for life through this unique Greenland record, we must fully exploit the spectrum of biosignatures available; these efforts must also be coupled with an understanding of the complex geologic history of the rocks hosting these signatures. The new methods employed here will eventually become applicable to other worlds when samples become available for study early in the 21st century.
INTRODUCTION
|
Figure 1. Environment of Hadean (>3800 Ma) Earth was more similar to than different from Earth of today. Liquid water and cycling of elements between geochemical reservoirs had already been established by 3.9 Ga. (Artwork by Don Dixon.) |
| If the presence of a stable liquid water veneer on Earth is necessary for the origin, evolution, and propagation of life (Fig. 1), then the best sources of information to understand this phenomenon are water-laid sediments preserved in the geologic record. Ancient (>3800 Ma) water-sculpted terrains have been recognized on Mars, and liquid water appears to exist beneath the icy crust of Jupiter's moon Europa. As long as liquid water, energy sources, and organic building blocks were present, life could have emerged on these worlds as well. The discovery of terrestrial marine sediments approaching 4 b.y. in age (Nutman et al., 1996) provides a remarkable opportunity to investigate possible environments from which life arose. Complicating this opportunity is their protracted metamorphic history; the oldest known sediments have been recrystallized under high-grade metamorphism (Griffin et al., 1980) and do not contain recognizable microfossils (Bridgwater et al., 1981). However, stable isotope fractionations of the bioessential elements (e.g., C, N, and S) produced by different metabolic styles can be preserved in ancient sediments (Schidlowski et al., 1983). This permits understanding of the development of early life inferred from chemical and isotopic information, rather than solely on interpretation of microfossil-like shapes (e.g., Schopf, 1993; McKay et al., 1996) (Fig. 2). | |
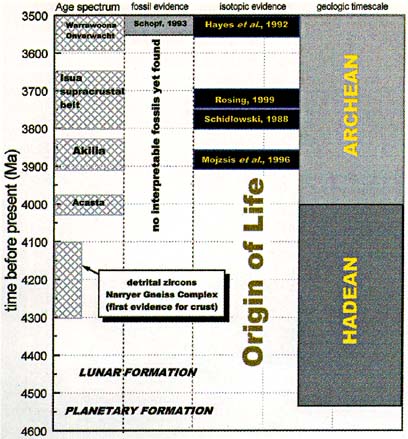
|
Figure 2. Timeline of early Earth history and recent advances made in investigating oldest biogeochemical records. Listed are ages for oldest rock sequences, and of isotopic evidence for life in oldest sediments. |
EARLY EARTH–SURFACE STATE
Early Earth was likely dominated by markedly different hydrospheric (e.g., lower pH, much higher [Fe2+]aq) and atmospheric (e.g., much lower pO2, much higher pCO2) conditions (Holland, 1984) and a tectonic style reflecting higher heat flow through the crust than at present. Several factors were unique to the early Archean surface, including a higher ultraviolet flux from a Sun 30% less luminous at 3800 Ma than today (Kuhn et al., 1989) and impact rates from asteroids and comets many orders of magnitude greater. Together, these conditions would presumably have restricted the number of suitable environments for life to emerge. The minimum ages of some of the oldest Greenland rocks (Nutman et al., 1996, 1997) appear to overlap in time with a period of intense impacts peaking at 3850 ± 100 Ma as recorded on the Moon (Ryder, 1990). Thermal and shock effects associated with the Late Heavy Bombardment era (Tera et al., 1974) are presumed to have rendered early Earth unsuitable for the emergence of life until after the massive bombardments ceased (e.g., Maher and Stevenson, 1988). During this bombardment era, conditions may have favored the survival of certain bacteria that survive (and even thrive) in environmental extremes of temperature, pressure, and pH before diversifying into wider ecological niches throughout the planet. Phylogenetic studies using highly conservative ribosomal RNA sequences reveal that the deepest branches of life derive from "heat-loving," or thermophilic, bacteria (Pace, 1997). Such organisms could have survived thermal assaults from giant impacts, especially if sequestered deep in the oceans or in rocks away from a destructive surface zone bathed both in the intense ultraviolet radiation of the early Sun and a rain of extraterrestrial debris ~4 b.y. ago. To better understand this early era, we have to unravel the timing of events from a heavily modified early Archean rock record.
EARLY ARCHEAN (>3500 Ma) HISTORY OF WEST GREENLAND
The diverse rock types present in the Isua district of West Greenland are all contained within extensive early Archean (3600–3900 Ma) gneisses dominantly of tonalitic-granodioritic composition (Black et al., 1971; Nutman et al., 1996) (Fig. 3). This multiply metamorphosed terrane, termed the Itsaq Gneiss Complex (Nutman et al., 1996), contains <10% volcano-sedimentary enclaves in the gneisses. The identification of water-laid rocks, such as banded iron-formation (BIF), metamorphosed chert, metapelites, rare graywackes, and pillow lava basalts, attests to
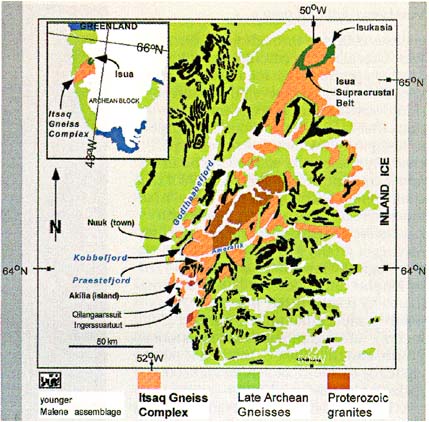 |
Figure 3. Generalized geologic map showing extent of early Archean (3770�3900 Ma) Itsaq Gneiss Complex in southern West Greenland (adapted from Nutman et al., 1984, 1996). Oldest sedimentary rocks and associated gneisses are along coast, represented by island of Akilia, and are intruded in places by Proterozoic Qorqut granites (Fig. 4). |
a hydrosphere with an already mature cycling of sediments by ~3900 Ma. The paleogeography of this sedimentary system remains poorly understood (e.g., oceanic arc-back-arc basin or deep abyssal basins), but the apparent lack of continentally derived sediment (Nutman et al., 1984) mitigates against the widespread presence of nearby exposed continental crust at the time of deposition.
Banded Iron-Formation >3850 Ma on Akilia Island
The oldest known sediment (Nutman et al., 1997), and the oldest known rock with evidence of biological processes active during time of deposition
(Mojzsis et al., 1996), is a layer ~3 m thick of BIF within a body of amphibolite on the southern tip of Akilia island, West Greenland (Fig. 4). This BIF on Akilia was chosen as the type locality for the Akilia association, a term used for volcano-sedimentary enclaves found throughout the Itsaq Gneiss Complex that are not part of the larger and better preserved Isua supracrustal belt (McGregor and Mason, 1977). More detailed field studies through the 1980s and 1990s, combined with geochronological work on the intruding gneisses in the coastal regions of southern West Greenland, demonstrated that the rocks around Godthåbsfjord (in the southwestern coastal area), including those of the Akilia association, contain older components
(Kinny, 1986) than most rocks in the Isua district, 150 km distant.
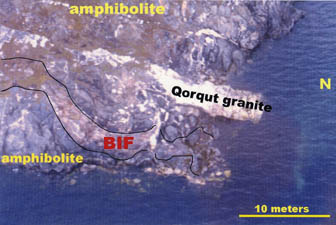 |
Figure 4. Aerial photograph of BIF locality on Akilia, looking east; sediments are bounded by amphibolites and intruded by gneissic sheets up to 3850 m.y. old (Fig. 5). |
On Akilia island itself, gneissic sheets crosscut the amphibolite enclave containing BIF and yield
U-Pb zircon ages as old as ca. 3850 Ma, interpreted as the age of crystallization of the magmatic protolith by Nutman et al. (1996, 1997). If this interpretation is correct, then the BIF is at least as old as 3850 Ma. Carbon isotopic evidence of bio-organic activity during deposition of the BIF sediments
(Mojzsis et al., 1996) would then suggest that the emergence of life on this planet occurred much earlier than previously thought (Hayes, 1996; Holland, 1997). This age for the Akilia island sediments is also significant because it would place their deposition simultaneous with the Late Heavy Bombardment of the Moon at 3800–3900 Ma, and coeval with the presence of abundant liquid water on the surface of Mars.
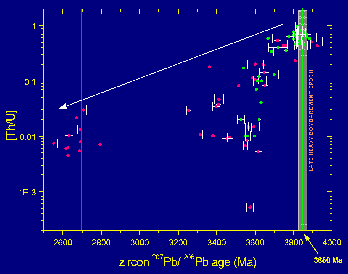 |
Figure 5. 207Pb/206Pb ion microprobe ages of zircons from tonalitic gneiss intruding amphibolite + banded iron-formation on Akilia island, West Greenland. What we interpret as the magmatic zircon population in these rocks (top right) has highest Th/U values and tightest spectrum of ages at 3850 � 10 Ma (data in green from Nutman et al., 1997; data in red from Mojzsis and Harrison, 1999). All other ages are for lower Th/U metamorphic zircons (arrow) that correlate with well-known metamorphic events in West Greenland at 3650 Ma and 2700 Ma (Baadsgaard et al., 1976; Griffin et al., 1980). |
Several of the Akilia gneisses that cut the BIF generally contain three zircon age populations: ca. 3850, 3650, and 2700 Ma (Fig. 5) (Mojzsis and Harrison, 1999; Whitehouse et al., 1999). One possible explanation for this zircon age distribution is that the ~2700 Ma grains are metamorphic in origin, the ~3650 Ma grains are igneous, and the oldest grains are xenocrysts inherited from an older rock. In the latter interpretation, the actual intrusive age of the tonalitic protolith of the gneiss would be only 3650 Ma. For example, Kamber and Moorbath (1998) and Whitehouse et al. (1999) argued that the ca. 3850 Ma ages were assimilated at about 750 °C from adjacent, zircon-bearing rocks. However, there is no identified candidate rock for the assimilant in the whole of the Itsaq Gneiss Complex. The granitoid protolith of the orthogneisses is characterized by relatively low Zr contents (~120 ppm) and high crystallization temperatures (>900 °C). The likelihood of strongly zircon undersaturated tonalitic-granodioritic melts (Harrison and Watson, 1983) intruding into zircon-poor rocks and preserving widespread inherited zircon is low (Watson, 1996). Kamber and Moorbath (1998) argued that the lack of 3850 Ma Pb-Pb ages in feldspars from gneisses collected throughout southern West Greenland specifically preclude a 3850 Ma protolith age for any rocks in West Greenland. However, their conclusion overstates the clarity of interpretations that can be drawn from Pb isotopes in feldspars. Peak metamorphism experienced by these rocks at 3650 Ma occurred under conditions (Griffin et al., 1980) that could permit exchange of primitive Pb isotopes in feldspar (Cherniak, 1992) with radiogenic Pb released from U-rich phases. McGregor (2000) pointed out that the petrologically unrelated samples analyzed by Kamber and Moorbath (1998) are dominated by relatively late phases of the gneiss complex that underwent partial melting under granulite facies metamorphism, and thus their data "do not preclude the existence of very old (greater than or equal to 3800 Ma) rocks" on Akilia island.
Existing evidence points to a 3850 Ma age for granitoids intruding BIF on Akilia island, but the great significance of this age for early terrestrial evolution leads us to continue our geochronological investigations in more detail. We are testing more directly the hypothesis that the 3850 Ma zircon ages are the result of xenocrystic contamination by determining in situ U-Pb ages of zircons included in preserved, early-crystallizing phases of the tonalite protolith. Also, the three-dimensional distribution of Pb/U revealed by isotopic depth profiling (e.g., Grove and Harrison, 1999) and applied to the oldest zircons helps us address whether zircon overgrowths reflect magmatic or metamorphic processes. If these rocks indeed provide a glimpse into pre–3800 Ma Hadean Earth, then the time of the emergence of the biosphere appears to have coincided with a period of intense bombardment in the inner solar system, as recorded on the Moon.
LATE HEAVY BOMBARDMENT OF THE MOON AND EARTH AT CA. 3850 MA
The surface of the Moon displays evidence of an intense bombardment that took place at some time between original crust formation and the outpourings of lava that form the dark mare plains. The oldest of the lunar mare are dated at ~3800 Ma (see review in McDougall and Harrison, 1999). Breccias from the lunar highlands generally yield radiometric ages between 3800 and 3900 Ma that are interpreted as reflecting impact resetting (Dalrymple and Ryder, 1996). The narrow distribution of ages and rapid transition from older, impact-dominated landforms to volcanic plains has generally been attributed to a cataclysmic spike in impacts at 3800–3900 Ma (Tera et al., 1974; Ryder, 1990). The present evidence is consistent with an abrupt termination of this intense activity coinciding with nearly simultaneous creation of the large Imbrium, Orientale (Fig. 6), and Schrödinger basins at 3850 Ma, or even as early as 3870 Ma (G. Ryder, personal communication). These timing details are important for at least two reasons: An age of 3850 Ma closely corresponds to that of the oldest sediments in West Greenland cited above, and that age has been taken as a temporal marker horizon for estimating the age of heavily cratered crust on Mars and Mercury.
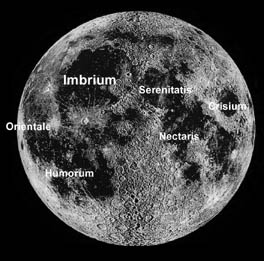 |
Figure 6. Telescopic mosaic of the front side of Moon shows dark mare basalt plains and much more rugged and heavily cratered highlands of higher albedo and more feldspathic composition. Highlands show ancient impact basins as well as craters. |
Consequences of Bombardments to Early Life
Highly energetic impacts would have been deleterious for near-surface emergent
ecosystems as conceptualized in the classic "Darwin Pond" hypothesis of the origin
of life. Impact "erosion" of planetary atmospheres and wholesale destruction of
hydrosphere and crust from the most massive infalling bodies are some of the effects
postulated to have rendered the surface zone truly Hadean in character before
3800 Ma (Sleep et al., 1989). However, if life originated in deep marine or crustal
nurseries (Shock et al., 1995) and continued to reside there until bombardment
ceased, events at the surface might have had little immediate consequence to survivability
of the biosphere.
To search for evidence of impacts in the geologic record, workers have taken advantage of the geochemistry of iridium. Earth's crust is highly depleted in Ir and other platinum-group metals relative to its mantle, which is itself depleted relative to chondritic meteorites by partitioning of Ir into the core. Iridium content of sediments is therefore used as a sensitive indicator of the presence of extraterrestrial debris (e.g., Alvarez et al., 1980). Sources of Ir in ancient sediments might represent interplanetary dust particles, micrometeorites, local- to world-spanning impacts, airbursts, and products of asteroid or comet showers. If depositional rates can be estimated, measurements of Ir in sediments can provide information on the accretion of extraterrestrial material during the time of deposition. On the basis of timing of the lunar bombardment and age of the oldest terrestrial sediments, the Akilia rocks might therefore be expected to preserve a signal of markedly higher flux according to their precise age correlation with the lunar bombardment record.
However, unless sedimentation rates were anomalously rapid (»1 mm/yr), thereby diluting a signal of intense impacts, the Ir contents for all West Greenland samples studied thus far are lower than that expected from simple models for the Late Heavy Bombardment (Anbar et al., 1999). This observation may be explained by the fact that the bombardment was not an era of continuous massive impacts. It was most likely dominated by quiet conditions punctuated by relatively infrequent episodes of extreme thermal shocks, none of which could have erased a deep-seated biosphere already in place. Furthermore, evidence for life from carbon isotopic distributions in rocks from the early Archean of Greenland would appear to indicate that life withstood the Late Heavy Bombardment, whatever its intensity, relatively unscathed.
CHEMICAL AND ISOTOPIC FOSSILS OF EARLY LIFE
On a global scale, kinetic isotope fractionations during enzymatic activity produce distinctive variations in stable isotope ratios of bioessential elements (e.g., carbon, nitrogen, and sulfur) in various reservoirs. A record of these fractionations can be obtained by measuring the isotopic ratios of these elements in bio-organic materials preserved in sediments. It has long been recognized (e.g., Thode et al., 1949; Wickman, 1952; Craig, 1953) that life systematically and significantly affects the isotopic composition of the biologically important elements and that these distinctive biosignatures can be preserved during diagenesis and moderate-grade metamorphism. After more than 50 years of investigations, geochemists now agree that postdepositional processes tend to obscure the original carbon isotopic signal of bio-organic material, but only toward values characteristic of inorganic carbon (Schidlowski, 1988; Kitchen and Valley, 1995; Des Marais, 1997). If this isotope record can survive very high grade metamorphism, then it becomes possible to analyze the oldest sediments for evidence of life (e.g., Mojzsis et al., 1996).
Carbon Isotopes
The primary metabolism of organisms on Earth that manufacture their own food (autotrophs)
imposes a kinetic isotope fractionation that discriminates against 13C in the
fixation of atmospheric CO2 (d13CPDB
= –7.9‰) and dissolved marine HCO–3 (d13C
= 0 ± 1‰). Because of the intrinsic lower reactivity of 13C
vs. 12C, enzymatic processes discriminate against CO2 during
carbon fixation (by the enzyme ribulose bisphosphate carboxylase in cyanobacteria
and plants; Park and Epstein, 1960). These mechanisms induce up to 5% differences
in isotopic composition of bio-organic carbon with respect to inorganic carbon
sources in the atmosphere and hydrosphere. The average isotopic composition of
Phanerozoic bio-organic matter (d13C = –27‰)
is far away from the inorganic field. While Horita and Berndt (1999) postulated
that the inorganic production of CH4 from serpentinization could create
a carbon isotope fractionation of similar magnitude, subsequent work suggests
that the evolved methane in these experiments derives exclusively from bio-organic
contaminants in the reactant olivine (McCollom and Seewald, 1999). No known abiotic
process mimics the large carbon isotopic signal of life in sediments.
Microbial methanogenesis is a widespread phenomenon whereby organic matter decomposes in anaerobic sediments and in the intestinal tracts of ruminants. The acetyl-CoA pathway — a metabolic system utilized by methanogens — is responsible for carbon isotope fractionations up to –40‰ from inorganic carbon (Preuß et al., 1989). Methanogens are a group of chemosynthetic (rather than photosynthetic) microbes that are among the most primitive life forms (Woese, 1987). Chemoautotrophes can produce biomass with d13C values as low as –60‰ (Summons et al., 1998). The recycling of carbon by these primitive microbes, and the isotopically light (d13C = –30‰ to –60‰) composition of their organic remains are considered diagnostic of contemporary microbial habitats characterized by extreme environments. These habitats (e.g., high salinity, high water temperature, or variable pH) are considered analogous to some of the earliest environments for life on Earth.
Analyses of bulk carbonaceous matter trapped within early Archean, Proterozoic, and younger sediments has demonstrated the ubiquity of isotopically light carbon (Schidlowski et al., 1979, 1983; Hayes et al., 1983; Des Marais, 1997; Rosing, 1999). These results are consistent with a bio-organic origin for sedimentary reduced carbon; yet, generalized concerns regarding later contamination or open-system behavior during metamorphism have been raised. While specific criticisms of this kind (e.g., Sano et al., 1999) can be shown to be unwarranted on geologic grounds alone (e.g., Mojzsis et al., 1999a), newly developed isotopic measurements permit textural and mineralogical preservation of the analyzed material and potentially transcend these two issues. Mojzsis et al. (1996) used a high-resolution ion microprobe to analyze extremely small (~10 mm) carbonaceous inclusions locked within mineralogic domains that had remained stable since diagenesis (Mojzsis et al., 1999a). Carbon isotopic values for carbonaceous inclusions in apatite from the greater than or equal to 3770 Ma Isua sediments (d13C = –30‰) and the greater than or equal to 3850 Ma BIF from Akilia island (d13C = –37‰) closely match those for similar rocks from a variety of younger, less metamorphosed rocks.
The simplest interpretation of the carbon isotopic data in Mojzsis et al. (1996) is that the organisms responsible for the light carbon signature in the oldest known terrestrial sediments were metabolically complex, perhaps comprising populations of phosphate-utilizing photoautotrophs and chemoautotrophs. These data may point to the presence of diverse photosynthesizing, methanogenic, and methylotrophic bacteria on Earth before 3850 Ma (Mojzsis and Arrhenius, 1998; Mojzsis et al., 1999b). Not only had life taken firm hold on Earth by the close of the Hadean era, but it also appears to have evolved far enough away from its origin to create an interpretable signature in carbon isotopes.
Our group at the University of California, Los Angeles, has undertaken study of the stable isotope composition of individual microfossils by the techniques described above; these studies indicate the opening of new vistas in the isotopic paleontology of ancient life forms where recognizable fossil microbes are preserved. Furthermore, the utility of sulfur isotope (32S,33S, 34S) measurements of individual sulfide minerals in ancient rocks (Greenwood et al., 1999) potentially provides information not only about primitive metabolic cycling of sulfur in the biosphere, but also of unique atmospheric fractionations and atmosphere-crust interactions early in Earth history.
ACKNOWLEDGMENTS
We thank Kevin McKeegan and Christopher Coath for ongoing scientific advice, and David Des Marais, Christopher House, and an anonymous reviewer for their constructive comments. We received research support from the National Science Foundation's Life in Extreme Environments (LexEn) and Earth Sciences Postdoctoral Fellowship programs, from NASA's Astrobiology Institute, from the Society of Economic Geologists Foundation (Hickok-Radford Award), and from the GSA research grants program; facility support came from the NSF Instrumentation and Facilities Program. This is a contribution to the Isua Multidisciplinary Research Project directed by P.W.U. Appel (GEUS).
Note by J. Schieber: Interpretation of above rocks as
metamorphosed sediments is under debate. An opposing view (short
summary) that would invalidate
above conclusions was published by
REFERENCES CITED
Alvarez, L.W., Alvarez, W., Asaro, F., and Michel, H.V., 1980, Extraterrestrial cause for the Cretaceous-Tertiary extinction: Science, v. 208, p. 1095–1108.
Anbar, A.D., Arnold, G.L., Mojzsis, S.J., and Zahnle, K.J., 1999, Extraterrestrial iridium and sediment accumulation on the Hadean Earth: 9th Annual V.M. Goldschmidt Conference, Abstract 7382: Houston, Lunar and Planetary Institute, LPI Contribution 971, (CD-ROM).
Baadsgaard, H., Lambert, R.S., and Krupicka, J., 1976, Mineral isotopic age relationships in the polymetamorphic Am"tsoq gneisses, Godthåb District, West Greenland: Geochimica et Cosmochimica Acta, v. 40, p. 513–527.
Black, L.P., Gale, N.H., Moorbath, S., Pankhurst, R.J., and McGregor, V.R., 1971, Isotope dating of very early Precambrian amphibolite facies gneisses from the Godthåb District, West Greenland: Grønlands Geologisk Undersølgelse Miscellaneous Paper 98, p. 245–259.
Bridgwater, D., Alaart, J.H., Schopf, J.W., Klein, C., Walter, M.R., Barghoorn, E.S., Strother, P., Knoll, A.H., and Gorman, B.E., 1981, Microfossil-like objects from the Archaean of Greenland: A cautionary note: Nature, v. 289, p. 51–53.
Cherniak, D.J., 1992, Diffusion of Pb in feldspar measured by Rutherford backscattering spectroscopy: Eos (Transactions, American Geophysical Union), v. 73, p. 641.
Craig, H., 1953, The geochemistry of the stable carbon isotopes: Geochimica et Cosmochimica Acta, v. 3, p. 53–92.
Dalrymple, G.B., and Ryder, G., 1996, 40Ar/39Ar age spectra of Apollo 17 highlands breccia by laser step-heating and the age of the Serenitatis basin: Journal of Geophysical Research, v. 101, p. 26,069–26,084.
Des Marais, D.J., 1997, Long-term evolution of the biogeochemical carbon cycle, in Banfield, J.F., and Nielsen, K.H., eds., Geomicrobiology: Interactions between microbes and minerals: Reviews in Mineralogy, v. 35, p. 429–448.
Greenwood, J.P., Mojzsis, S.J., Coath, C.D., and Wasson, J.T., 1999, Measurements of 32S, 33S, 34S in AH84001 and Nakhla sulfides by multicollector-SIMS: Implications for crustal-atmospheric exchange and biogenic activity on Mars: 9th Annual V.M. Goldschmidt Conference, Abstract 7601: Houston, Lunar and Planetary Institute, LPI Contribution 971, (CD-ROM).
Griffin, W.L., McGregor, V.R., Nutman, A.P., Taylor, P.N., and Bridgwater, D., 1980, Early Archaean granulite facies metamorphism south of Ameralik, West Greenland: Earth and Planetary Science Letters, v. 50, p. 59–74.
Grove, M. and Harrison, T.M., 1999, Monazite Th-Pb age depth profiling: Geology, v. 27, p. 487–490.
Harrison, T.M., and Watson, E.B., 1983, Kinetics of zircon dissolution and zirconium diffusion in granitic melts of variable water content: Contributions to Mineralogy and Petrology, v. 84, p. 66–72.
Hayes, J.M., 1996, The earliest memories of life on Earth: Nature, v. 384, p. 21–22.
Hayes, J.M., Kaplan, I.R., and Wedeking, K.W., 1983, Precambrian organic chemistry; preservation of the record, in Schopf, J.W., ed., Earth's earliest biosphere-Its origin and evolution: Princeton, New Jersey, Princeton University Press, p. 53–134.
Hayes, J.M., Des Marais, D., Lambert, H., Strauss, H., and Summons, R.E., 1992, Proterozoic biogeochemistry, in Schopf, J.W., and Klein, C., eds., The Proterozoic biosphere: New York, Cambridge University Press, p. 81–134.
Holland, H.D., 1984, The chemical evolution of the atmosphere and oceans: Princeton, New Jersey, Princeton University Press, 582 p.
Holland, H.D., 1997, Evidence for life on Earth more than 3,850 million years ago: Science, v. 275, p. 38–39.
Horita, J., and Berndt, M.E., 1999, Abiogenic methane formation and isotopic fractionation under hydrothermal conditions: Science, v. 285, p. 1055–1057.
Kamber, B.S., and Moorbath, S., 1998, Initial Pb of the Amîtsoq gneiss revisited: Implication for the timing of the early Archaean crustal evolution in West Greenland: Chemical Geology, v. 150, p. 19–41.
Kinny, P., 1986, 3820 Ma zircons from a tonalitic Amîtsoq gneiss in the Godthåb district of southern West Greenland: Earth and Planetary Science Letters, v. 79, p. 337–347.
Kitchen, N.E., and Valley, J.W., 1995, Carbon isotope thermometry in marbles of the Adirondack Mountains, New York: Journal of Metamorphic Geology, v. 13, p. 577–594.
Kuhn, W.R., Walker, J.C.G., and Marshall, H.G., 1989, The effect on Earth's surface temperature from variations in rotation rate, continent formation, luminosity, and carbon dioxide: Journal of Geophysical Research, v. 94, p. 11,129–11,136.
Maher, K.A., and Stevenson, D. J., 1988, Impact frustration and the origin of life: Nature, v. 331, p. 612–614.
McCollom, T.M., and Seewald, J.S., 1999, Rapid equilibration of CO2 and formate under hydrothermal conditions (with a comment on the abiotic synthesis of hydrocarbons during serpentinization): Geological Society of America Abstracts with Programs, v. 31, no. 7, p. A431.
McDougall, I., and Harrison, T.M., 1999, Geochronology and thermochronology by the 40Ar/39Ar method, 2nd ed.: New York, Oxford University Press, 269 p.
McGregor, V.R., 2000, Age significance of U-Th-Pb zircon data from early Archaean rocks of West Greenland — A reassessment based on combined ion-microprobe and imaging studies — Comment: Chemical Geology (in press).
McGregor, V.R., and Mason, B., 1977, Petrogenesis and geochemistry of metabasaltic and metasedimentary enclaves in the Amîtsoq gneisses, West Greenland: American Mineralogist, v. 62, p. 887–904.
McKay, D.S., Gibson, E.K., Thomas-Keprta, K.L., Vali, H., Romanek, C.S., Clemett, S.J., Chillier, X.D.F., Maeschling, C.R., and Zare, R.N., 1996, Search for past life on Mars: Possible relic biogenic activity in martian meteorite ALH84001: Science, v. 273, p. 924–930.
Mojzsis, S.J., and Arrhenius, G., 1998, Phosphate and carbon on Mars: Exobiological implications and sample return considerations: Journal of Geophysical Research, v. 103, p. 28,495–28,511.
Mojzsis, S.J., and Harrison, T.M., 1999, Geochronological studies of the oldest known marine sediments: 9th Annual V.M. Goldschmidt Conference, Abstract 7602: Houston, Lunar and Planetary Institute, LPI Contribution 971 (CD-ROM).
Mojzsis, S.J., Arrhenius, G., McKeegan, K.D., Harrison, T.M., Nutman, A.P., and Friend, C.R.L., 1996, Evidence for life on Earth before 3,800 million years ago: Nature, v. 384, p. 55–59.
Mojzsis, S.J., Harrison, T.M., Arrhenius, G., McKeegan, K.D., and Grove, M., 1999a, Origin of life from apatite dating? — Reply: Nature, v. 400, p. 127–128.
Mojzsis, S.J., Krishnamurthy, R., and Arrhenius, G., 1999b, Before RNA and after — Geophysical and geochemical constraints on molecular evolution, in Gesteland, R., et al., eds., RNA world, 2nd ed.: Cold Spring Harbor, New York, Cold Spring Harbor Laboratory Press, p. 1–49.
Nutman, A.P., Allaart, J.H., Bridgwater, D., Dimroth, E., and Rosing, M., 1984, Stratigraphic and geochemical evidence for the depositional environment of the early Archaean Isua supracrustal belt, southern West Greenland: Precambrian Research, v. 25, p. 365–396.
Nutman, A.P., McGregor, V.R., Friend, C.R.L., Bennett, V.C., and Kinny, P.D., 1996, The Itsaq Gneiss Complex of southern West Greenland: The world's most extensive record of early crustal evolution (3900–3600 Ma): Precambrian Research, v. 78, p. 1–39.
Nutman, A.P., Mojzsis, S.J., and Friend, C.R.L., 1997, Recognition of >3850 Ma water-lain sediments and their significance for the early Archaean Earth: Geochimica et Cosmochimica Acta, v. 61, p. 2475–2484.
Pace, N.R., 1997, A molecular view of microbial diversity and the biosphere: Science, v. 276, p. 734–740.
Park, R., and Epstein, S., 1960, Carbon isotope fractionation during photosynthesis: Geochimica et Cosmochimica Acta, v. 21, p. 110–126.
Preuß, A., Schauder, R., Fuchs, G., and Stichler, W., 1989, Carbon isotope fractionation by autotrophic bacteria with three different CO2 fixation pathways: Zeitschrift für Naturforschung, v. 44, p. 397–402.
Rosing, M.T., 1999, 13C-depleted carbon microparticles in >3700 Ma seafloor sedimentary rocks from West Greenland: Science, v. 283, p. 674–676.
Ryder, G., 1990, Lunar samples, lunar accretion and the early bombardment of the Moon: Eos (Transactions, American Geophysical Union), v. 71, p. 322–323.
Sano, Y., Terada, K., Takahashi, Y., and Nutman, A.P., 1999, Origin of life from apatite dating?: Nature, v. 400, p. 127.
Schidlowski, M., 1988, A 3,800 million-year-old record of life from carbon in sedimentary rocks: Nature, v. 333, p. 313–318.
Schidlowski, M., Appel, P.W.U., Eichmann, R., and Junge, C.E., 1979, Carbon isotope geochemistry of the 3.7 × 109 yr old Isua sediments, West Greenland: Implications for the Archaean carbon and oxygen cycles: Geochimica et Cosmochimica Acta, v. 43, p. 189–199.
Schidlowski, M., Hayes, J.M., and Kaplan, I.R., 1983, Isotopic inferences of ancient biochemistries: Carbon, sulfur, hydrogen and nitrogen, in Schopf, J.W., ed., Earth's earliest biosphere — Its origin and evolution: Princeton, New Jersey, Princeton University Press, p. 149–186.
Schopf, J.W., 1993, Microfossils of the early Archean Apex Chert: New evidence for the antiquity of life: Science, v. 260, p. 640–646.
Shock, E.L., McCollom, T., and Schulte, M.D., 1995, Geochemical constraints on chemolithoautotrophic reactions in hydrothermal systems: Origin of Life and Evolution of the Biosphere, v. 25, p. 141–159.
Sleep, N.H., Zahnle, K.J., Kasting, J.F., and Morowitz, H., 1989, Annihilation of ecosystems by large asteroid impacts on the early Earth: Nature, v. 342, p. 139–142.
Summons, R.E., Franzmann, P.D., and Nichols, P.D., 1998, Carbon isotopic fractionation associated with methylotrophic methanogenesis: Organic Geochemistry, v. 28, p. 465–475.
Tera, F., Papanastassiou, D.A., and Wasserburg, G.J., 1974, Isotopic evidence for a terminal lunar cataclysm: Earth and Planetary Science Letters, v. 22, p. 1–21.
Thode, H.G., Macnamara, J., and Collins, C.B., 1949, Natural variations in the isotopic composition of sulfur and their significance: Canadian Journal of Research, v. 27B, p. 361.
Watson, E.B., 1996, Dissolution, growth and survival of zircons during crustal fusion: Kinetic principles, geological models, and implications for isotope inheritance: Royal Society of Edinburgh Transactions, v. 87, p. 43–57.
Whitehouse, M., Kamber, B.S., and Moorbath, S., 1999, Age significance of U-Th-Pb zircon data from early Archaean rocks of West Greenland — A reassessment based on combined ion-microprobe and imaging studies: Chemical Geology, v. 160, p. 201–224.
Wickman, F.E., 1952, Variation in the relative abundance of carbon isotopes in plants: Geochimica et Cosmochimica Acta, v. 2, p. 243–254.
Woese, C.R., 1987, Bacterial evolution: Microbial Research, v. 51, p. 221–271.